Lífsgerving kólesteróls og lífefnafræði þess - Sykursýki
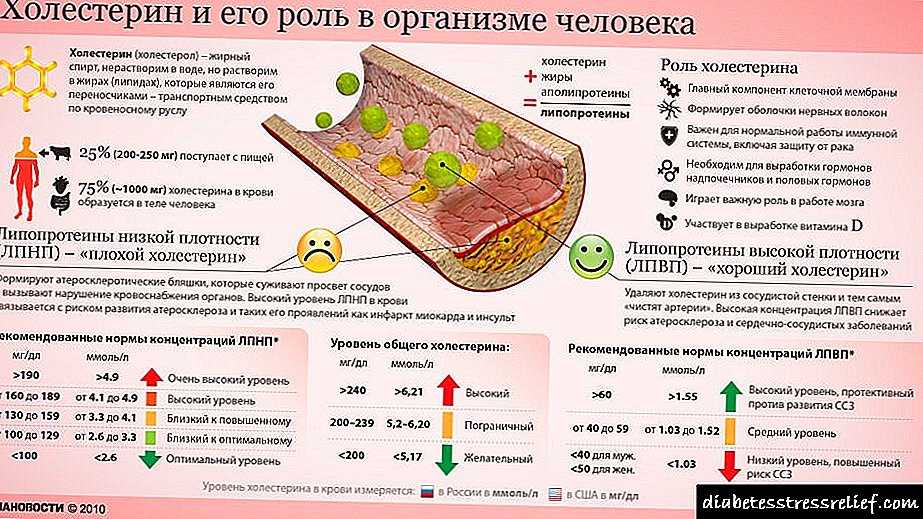
Án efa er kólesteról þekktasta lípíðið fyrir almenning, það er alræmt vegna mikillar fylgni milli hás kólesteróls í blóði og tíðni hjarta- og æðasjúkdóma hjá mönnum. Minni athygli hefur verið vakin á mikilvægu hlutverki kólesteróls sem hluti frumuhimna og sem undanfari sterahormóna og gallsýra. Kólesteról er nauðsynlegt fyrir mörg dýr, þar með talið menn, en nærvera þess í fæðu spendýra er valkvæð - líkamsfrumur geta sjálfar myndað það frá einföldum undanfari.
Uppbygging þessa 27 kolefnis efnasambands bendir til flókinnar leiðar til að mynda myndun þess, en öll kolefnisatóm þess er veitt af einum undanfari - asetati. Ísópren blokkir - mikilvægustu milliefnin frá asetati til kólesteróls, þau eru undanfara margra náttúrulegra lípíða, og aðferðir sem ísóprenblokkir eru fjölliðaðir eru svipaðir á öllum efnaskiptaferlum.
Við byrjum á því að skoða helstu stig í leiðinni til að mynda kólesteról úr asetati, ræðum síðan flutning kólesteróls í gegnum blóðrásina, frásog þess með frumum, eðlilega stjórnun á nýmyndun kólesteróls og reglugerð í tilfellum skertra frásogs eða flutninga. Þá skoðum við önnur efni sem koma frá kólesteróli, svo sem gallsýrum og sterahormónum. Að lokum, lýsing á líffræðilegum leiðum til myndunar margra efnasambanda - afleiður ísóprenblokka, þar sem algeng fyrstu stig eru með kólesterólmyndun, munu sýna fram á ótrúlega fjölhæfni ísóprenóíð þéttingar í lífmyndun.
Kólesteról er framleitt úr asetýl-CoA í fjórum stigum
Kólesteról, eins og langkeðju fitusýrur, er framleitt úr asetýl-CoA, en samsetningarmynstrið er allt öðruvísi. Í fyrstu tilraunum var asetati merkt með 14C annað hvort við metýl eða karboxýl kolefnisatóm bætt við fóður. Byggt á dreifingu merkimiða í kólesteróli sem einangrað var frá tveimur hópum dýra (mynd 21-32) var ensímstigum myndun kólesteróls lýst.
Mynd. 21-32. Uppruni kolefnisatóms í kólesteróli. Auðkennd við tilraunir með því að nota geislavirkt asetat merkt með metýl kolefni (svart) eða karboxýl kolefni (rautt). Í þéttu skipulaginu eru hringir táknaðir með bókstöfunum A til D.

Myndunin fer fram í fjórum stigum, eins og sýnt er á mynd. 21-33: (1) þétting þriggja asetatsleifa til að mynda sex kolefnis milliefni mevalónats, (2) umbreytingu mevalonats í virkjaðar ísóprenblokkir, (3) fjölliðun sex fimm kolefnis ísópreneininga til að mynda 30 kolefnis línulegt skvalen, (4) hringmyndun skvalen til að mynda fjórir hringir í sterakjarnanum og síðan röð breytinga (oxun, fjarlæging eða flutningur metýlhópa) við myndun kólesteróls.
Mynd. 21-33. Almenn mynd af nýmyndun kólesteróls. Fjallað er um fjögur stig myndunar í textanum. Ísóprenblokkir í skvalen eru merktir með rauðum strikuðum línum.

Stig (1). Tilmyndun mevalonats frá asetati. Fyrsta stigið í nýmyndun kólesteróls leiðir til myndunar milliefnis mevalonate (Mynd 21-34). Þessar tvær asetýl CoA sameindir þéttast til að gefa acetóasetýl CoA, sem þéttist við þriðju asetýl CoA sameindina til að mynda sex kolefnis efnasamband ß-hýdroxý-ß-metýlglutaryl-CoA (HM G-COA). Þessi tvö fyrstu viðbrögð eru hvötuð thiolase og NM G-COA synthase, í sömu röð. Frumueyðandi NM G-CoA synthase Þessi efnaskiptaferill er frábrugðinn hvatbera ísóensíminu, sem hvatar myndun NM G-COA við myndun ketónlíkama (sjá mynd 17-18).
Mynd. 21-34. Myndun mevalonats úr asetýl-CoA. Uppruni C-1 og C-2 mevalonats frá asetýl-CoA er auðkenndur með bleiku.

Þriðja viðbrögðin takmarka hraðann á öllu ferlinu. Í henni er NM G -CoA minnkað í mevalonat, sem hver af tveimur NА D PH sameindum veitir tvær rafeindir. HMG-CoA reductase - samþætt himnurprótein af sléttu ER, það þjónar, eins og við munum sjá síðar, sem aðalatriðið í stjórnun á efnaskiptaferli kólesterólmyndunar.
Stig (2). Umbreytingu mevalonats í tvö virkjuð ísópren. Á næsta stigi nýmyndunar kólesteróls eru þrír fosfathópar fluttir frá ATP sameindum til mevalonats (mynd 21-35). Fosfatið sem er bundið við hýdroxýlhópinn við C-3 mevalonat í millistiginu 3-fosfó-5-pýrófosfómevalónat er góður skilningshópur, í næsta skrefi fara bæði þessi fosföt og aðliggjandi karboxýl hópur, og myndar tvöfalt tengi í fimm kolefnisafurðinni ∆ 3 -ísópentenýl pýrófosfat. Þetta er fyrsta af tveimur virkjuðum ísóprenum - aðal þátttakendur í nýmyndun kólesteróls. Líkamsbreyting á Δ 3-ísópenentenýlpýrófosfat gefur annað virkjað ísópren dímetýlallyl pýrofosfat. Nýmyndun ísópentenýl pýrofosfats í umfrymi plöntufrumna fer fram samkvæmt þeirri leið sem lýst er hér. Plöntuklórplasts og margar bakteríur nota þó leið óháð mevalonate. Þessi valleið er ekki að finna hjá dýrum, svo hún er aðlaðandi þegar ný sýklalyf eru búin til.
Mynd. 21-35. Umbreyting mevalonats í virkjaðar ísóprenblokkir. Sex virkjuðu einingarnar myndast saman squalen (sjá mynd 21-36). Horfandi hópar 3-fosfó-5-pýrofosfómevalónats eru auðkenndir með bleiku. Í fermetra sviga er ímyndaður millistig.

Stig (3). Þétting sex virkjaðra ísópren eininga til að mynda skvalen. Ísópentenýl pýrofosfat og dímetýlallyl pýrofosfat fara nú í þéttingu frá höfuð til hala, þar sem einn pýrofosfat hópur hreyfist og 10 kolefnis keðja myndast - geranyl pýrofosfat (Mynd 21-36). (Pýrofosfat festist við höfuðið.) Geranyl pýrofosfat gengst undir eftirfarandi höfuð-í-hala þéttingu með ísópenentenýl pýrofosfat, og 15 kolefnis milliefni myndast farnesýl pýrófosfat. Að lokum sameina tvær sameindir farnesýl pýrofosfats „höfuð við höfuð“, báðar fosfathóparnir eru fjarlægðir - myndaðir skvalen.
Mynd. 21-36. Squalene myndun. Skúfalbygging sem inniheldur 30 kolefnisatóm á sér stað við þéttingu í röð sem er virkjuð með ísópren (fimm kolefnis) blokkum.

Algengu nöfnin á þessum milliefnum koma frá nöfnum upprunanna sem þau voru fyrst einangruð frá. Geraniol, hluti af rósolíu, hefur geranium bragð, og farnesól, sem er að finna í litum acacia farnesa, er lilja í dalnum ilm. Mörg náttúruleg plöntulykt tilheyrir efnasamböndum sem byggð eru úr ísóprenblokkum. Squalene, fyrst einangrað úr hákarlalifur (Squalus tegundir), samanstendur af 30 kolefnisatómum: 24 atómum í aðalkeðjunni og sex atómum í málmsuppbótarefnunum.
Stig (4). Umbreyting squalen í fjóra hringi í stera kjarna. Á mynd. 21-37 sést greinilega að uppbygging squalene keðjunnar og steróla - hringlaga. Öll steról eru með fjóra þéttaða hringa sem mynda stera kjarnann og allir eru þeir alkóhól með hýdroxýl hóp við C-3 atómið, þar með enska nafnið steról. Undir aðgerð squalene monooxygenase einu súrefnisatómi frá O er bætt við lok squalene keðjunnar 2 og myndast epoxíð. Þetta ensím er annar oxíðasi með blandaða virkni (bæta við 21-1), NADPH dregur úr öðru súrefnisatómi úr O 2 til H2 O. Tvöföld bönd vöru skvalen-2,3-epoxíð raðað þannig að ótrúlega stöðug viðbrögð geta breytt keðju skvalenoxíðs í hringlaga byggingu. Í dýrafrumum leiðir þessi hringrás til myndunar lanósteról sem inniheldur fjóra hringi sem eru einkennandi fyrir stera kjarna. Fyrir vikið er lanósteróli breytt í kólesteról með röð um það bil 20 viðbragða, sem felur í sér flæði sumra málmhópa og brottnám annarra. Lýsingin á þessari mögnuðu braut líffræðilegrar myndunar, sem er ein sú erfiðasta meðal þekktra, var gerð af Conrad Bloch, Theodore Linen, John Cornfort og George Popiak seint á sjötta áratugnum.
Mynd. 21-37. Hringlokun breytir línulegri skvalen í þéttan stera kjarna. Fyrsta stigið er hvatað með oxíðasa með blönduðu hlutverki (monooxygenase), sem samsöfnun er N AD PH. Varan er epoxíð, sem á næsta stigi hjólar til að mynda stera kjarna. Lokaafurð þessara viðbragða í dýrafrumum er kólesteról; í öðrum lífverum myndast steról sem eru aðeins frábrugðin því.


Kólesteról er steról sem einkennir dýrafrumur, plöntur, sveppir og protists framleiða önnur mjög svipuð steról.
Þeir nota sömu nýmyndunarleið yfir í skvalen-2,3-epoxíð en síðan víkja stígarnir örlítið og önnur steról myndast, svo sem sigmasterol í mörgum plöntum og ergósteról í sveppum (mynd 21-37).
Dæmi 21-1 Orkukostnaður fyrir Squalene nýmyndun
Hver er orkukostnaðurinn (gefinn upp sem ATP sameindir) við myndun einnar skvalen sameindar?
Lausn. Við myndun skvalens úr asetýl-CoA er ATP eingöngu varið á stiginu þegar mevalonat er breytt í virkan ísópren skvalen undanfara. Sex virkjaðar ísópren sameindir eru nauðsynlegar til að byggja upp squalen sameind og þrjár ATP sameindir eru nauðsynlegar til að framleiða hverja virku sameind. Alls er 18 ATP sameindum varið í nýmyndun einnar rúllu sameindar.
Efnasambönd af kólesteróli í líkamanum
Hjá hryggdýrum er mikið magn af kólesteróli búið til í lifur. Sumt af kólesterólinu sem þar er myndað er fellt inn í himnur lifrarfrumna, en það er aðallega flutt út í einni af þremur gerðum þess: gallkólesteróli (gall), gallsýrum eða kólesterólesterum. Gallsýrur og sölt þeirra eru vatnssæknar afleiður kólesteróls, sem eru búnir til í lifur og stuðla að meltingu lípíða (sjá mynd 17-1). Estrarar kólesteróls myndast í lifur með verkun acýl-CoA-kólesteról-asýltransferasa (ACAT). Þetta ensím hvetur flutning fitusýruleifar úr kóensími A til hýdroxýlhópsins á kólesteróli (mynd 21-38) og breytir kólesteróli í vatnsfælna formið. Kólesterólesterar í seyttum lípóprótein agnum eru fluttir í aðra vefi með kólesteróli eða geymdir í lifur.
Mynd. 21-38. Nýmyndun kólesterólesterna. Etherification gerir kólesteról enn vatnsfælin form til geymslu og flutninga.

Kólesteról er nauðsynlegt fyrir alla vefi vaxandi dýraveru til nýmyndunar á himnur og sum líffæri (til dæmis nýrnahetturnar og kynkirtlar) nota kólesteról sem undanfara sterahormóna (þetta verður fjallað hér að neðan). Kólesteról er einnig undanfari D-vítamíns (sjá mynd 10-20, v. 1).
Kólesteról og önnur lípíð eru með lípóprótein í plasma
Kólesteról og kólesterólesterar, eins og tríasýlglýseról og fosfólípíð, eru nánast óleysanleg í vatni, en þau verða að fara frá vefnum sem þau voru búin til í vefina þar sem þeir verða geymdir eða neyttir. Þau eru borin af blóðrásinni í formi blóðfituprótein í blóði - macromolecular fléttur af sérstökum burðarpróteinum (apolipoproteins) með fosfólípíðum, kólesteróli, kólesterólesterum og tríasýlglýserólum sem eru til staðar í þessum fléttum í ýmsum samsetningum.
Apólípróprótein („apo“ vísar til fitusnauðu próteinsins sjálfrar) sameina við lípíð og mynda nokkur brot af lípóprótein agnum - kúlulaga fléttur með vatnsfælnum lípíðum í miðjunni og vatnsæknum amínósýrukeðjum á yfirborðinu (mynd 21-39, a). Með ýmsum samsetningum af lípíðum og próteinum myndast agnir með mismunandi þéttleika - frá kýlómíkrónum til háþéttni fitupróteina. Hægt er að aðskilja þessar agnir með ofurskiljun (tafla 21-1) og sjá sjónrænt með rafeindasmásjá (mynd 21-39, b). Hvert brot af lípópróteinum hefur sérstaka virkni sem ræðst af myndunarstað, lípíðsamsetningu og innihaldi apólíprópróteina. Að minnsta kosti 10 mismunandi apólipóprótein fundust í blóðvökva manna (tafla 21-2), sem eru mismunandi að stærð, viðbrögð við sérstökum mótefnum og einkennandi dreifing í mismunandi flokkum af lípópróteinum. Þessir próteindir hlutar virka sem merkjaefni sem beina lípópróteinum að sérstökum vefjum eða virkja ensím sem virka á lípóprótein.
Tafla 21-1. Fitupróteinprótein úr mönnum
Samsetning (massahlutfall,%)
r = 513.000). Agni LDL inniheldur kjarna sem er um það bil 1.500 sameindir af kólesterólesterum, umhverfis kjarnann er skel með 500 sameindir kólesteróls, 800 sameindir af fosfólípíðum og ein sameind apoB-100. b - fjórir flokkar lípópróteina, sjáanlegir með rafeindasmásjá (eftir birtingu neikvæðra). Réttsælis, frá efri vinstri mynd: kylómíkrónur - með þvermál 50 til 200 nm, PL O NP - frá 28 til 70 nm, HDL - frá 8 til 11 nm, og LDL - frá 20 til 55 nm. Eiginleikar fitupróteina eru gefnir í töflu. 21-2.

Kýlómíkron, sem vísað er til í Sec. 17, færa mat triacylglycerols frá þörmum yfir í aðra vefi. Þetta eru stærstu lípópróteinin, þau eru með lægsta þéttleika og hæsta hlutfallslega innihald triacylglycerols (sjá mynd 17-2). Chylomicrons eru búnir til í ER í þekjufrumum sem fóðra smáþörminn, fara síðan í gegnum eitilkerfið og fara í blóðrásina í gegnum vinstri subclavian bláæð. Chylomicron apolipoproteins innihalda apoB-48 (einstakt fyrir þennan flokk lípópróteina), apoE og apoC-II (tafla 21-2). AroC-II virkjar lípóprótein lípasa í háræðum fituvefja, hjarta, beinvöðva og mjólkandi brjóstkirtla, sem tryggir flæði frjálsra fitusýra inn í þessa vefi. Þannig flytja chylomicrons matar fitusýrur yfir í vefi, þar sem þeir verða neyttir eða geymdir sem eldsneyti (mynd 21-40). Chylomicron leifar (aðallega leystar frá triacylglycerols, en samt innihalda kólesteról, apoE og apoB-48) eru fluttar um blóðrásina til lifrarinnar. Í lifur bindast viðtakar apoE sem er að finna í chylomicron leifum og miðla frásogi þeirra með endocytosis. Í lifrarfrumum losa þessar leifar kólesterólið sem þau innihalda og eyðileggjast í lýsósómum.
Tafla 21-2. Humal plasma lípóprótein apólípóprótein
Aðgerð (ef þekkt)
Virkar L CAT, hefur samskipti við ABC flutningsaðila
Hindrar L CAT
Virkar L CAT, kólesterólflutning / úthreinsun
Binst við LDL viðtaka
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Chylomicrons, VLDL, HDL
Byrjar úthreinsun VLDL og chylomicron leifa
Þegar matur inniheldur fleiri fitusýrur en nú er hægt að nota sem eldsneyti, breytast þær í triacylglycerols í lifur, sem mynda brot með sérstökum apólipópróteinum mjög lítill þéttleiki lípóprótein (VLDL). Óhóflegum kolvetnum í lifur er einnig hægt að breyta í triacylglycerols og flutt út sem VLDL (mynd 21-40, a).Auk triacylglycerols, inniheldur VLDL brotið ákveðið magn af kólesteróli og kólesterólesterum, svo og apoB-100, apoC-1, apoC-II, apoC III og apoE (tafla 21-2). Þessi lípóprótein eru einnig flutt með blóði frá lifur í vöðva og fituvef, þar sem, eftir að lípóprótein lípasi er virkjaður með apo-C II, eru frjálsar fitusýrur losaðar frá triacylglycerols í VLDL brotinu. Adipocytes ná ókeypis fitusýrum, breyta þeim aftur í triacylglycerols, sem eru geymdar í þessum frumum í formi fituinnlags (dropar), myocytes, þvert á móti, oxa fitusýrur strax til að mynda orku. Flestar VLDL leifar eru fjarlægðar úr blóðrásinni með lifrarfrumum. Upptöku þeirra, svipað og frásog chylomicrons, er miðlað af viðtökum og fer eftir nærveru apoE í VLDL leifum (í viðbót. 21-2 er sambandinu milli apoE og Alzheimers sjúkdóms lýst).
Mynd. 21-40. Fituprótein og lípíðflutningur, og - lípíð eru flutt um blóðrásina í formi lípópróteina, sem eru sameinuð í nokkur brot með mismunandi aðgerðir og mismunandi samsetningu próteina og fituefna (tab. 21-1, 21-2) og samsvarar þéttleika þessara hluta. Matarlípíð eru samsöfnuð í kýlómíkrón, flest triasýlglýserólin sem eru í þeim losa með lípóprótein lípasa í fitu og vöðvavef í háræðunum. Chylomicron leifar (sem innihalda aðallega prótein og kólesteról) eru teknar með lifrarfrumum. Innræn lípíð og kólesteról úr lifur eru borin í fitu og vöðvavef í formi VLDL. Losun lípíða frá VLDL (ásamt tapi nokkurra apólipópróteina) breytir smám saman VLDLP í LDL, sem skilar kólesteróli í vefjum utan lifrar eða skilar því í lifur. Lifrin fangar leifar VLDL, LDL og leifar chylomicrons með viðtaka-miðluðum endocytosis. Umfram kólesteról í vefjum utan lifrar er flutt aftur í lifur í formi LDL. Í lifur breytist hluti kólesterólsins í gallsölt. b - blóðsýni í blóðinu tekið eftir hungri (vinstri) og eftir að hafa borðað mat með hátt fituinnihald (til hægri). Kýlómíkrónur sem myndast með því að borða feitan mat gefa blóðvökva ytri líkingu mjólkur.

Með tapi á triacylglycerols er hluta VLDL breytt í VLDL leifar, einnig kallaðar millistærð lípóprótein (VLDL), frekari fjarlæging á triacylglycerols úr VLDL gefur lípóprótein með lágum þéttleika (LDL) (flipi. 21-1). LDL-hlutinn, sem er mjög ríkur í kólesteróli og kólesterólesterum, og inniheldur einnig apoB-100, flytur kólesteról yfir í lifrarvef sem bera sérstaka viðtaka sem þekkja apoB-100 á plasmahimnum sínum. Þessir viðtakar miðla upptöku kólesteróls og kólesterólesterna (eins og lýst er hér að neðan).
Viðbót 21-2.ApoE samsætur ákvarða tíðni Alzheimerssjúkdóms
Hjá mannfjölda eru þrjú þekkt afbrigði (þrír samsætur) af geninu sem kóðar apólípróprótein E. Af apoE samsætunum er APOEZ samsætan algengust hjá mönnum (u.þ.b. 78%), APOE4 samsætin eru 15 og 7%, talið í sömu röð. APOE4 samsætan er sérstaklega einkennandi fyrir fólk með Alzheimerssjúkdóm og þessi tengsl gera kleift að spá fyrir um tíðni sjúkdómsins með miklum líkum. Fólk sem hefur erft APOE4 er í mikilli hættu á að fá seint Alzheimerssjúkdóm. Fólk sem er arfhrein fyrir APOE4 er 16 sinnum líklegra til að þróa sjúkdóminn, meðalaldur þeirra sem veikjast er um það bil 70 ára. Hjá fólki sem erfir tvö eintök af AROEZ er þvert á móti meðalaldur Alzheimerssjúkdóms yfir 90 ár.
Sameindargrundvöllur fyrir tengsl apoE4 og Alzheimerssjúkdóms er ennþá óþekktur. Að auki er enn óljóst hvernig apoE4 getur haft áhrif á vöxt amyloid snúra, sem eru greinilega undirrót Alzheimerssjúkdóms (sjá mynd 4-31, v. 1). Forsendur beina sjónum að mögulegu hlutverki apoE við að koma á stöðugleika uppbyggingar frumuþvottar taugafrumna. ApoE2 og apoEZ próteinin bindast fjölda próteina sem tengjast örtöflum taugafrumna en apoE4 binst ekki. Þetta getur flýtt fyrir dauða taugafrumna. Hvað sem þetta fyrirkomulag kann að reynast, þessar athuganir vona að auka skilning okkar á líffræðilegum hlutum apólíprópróteina.
Fjórða tegund lípópróteina - háþéttni fituprótein (HDL), þetta brot myndast í lifur og smáþörmum í formi smápróteinríkra agna sem innihalda tiltölulega lítið kólesteról og alveg laust við kólesterólester (mynd 21-40). HDL brotið inniheldur apoA-I, apoC-I, apoC-II og önnur apolipoproteins (tafla 21-2), svo og lesitín-kólesteról-asýltransferasa (LC AT), sem hvatar myndun kólesterólesterna úr lesitíni (fosfatidýlkólíni) og kólesteróli (mynd 21-41). L CAT á yfirborði nýstofnaðra HDL agna breytir kýlómíkrón kólesteróli og fosfatidýlkólíni og VLDL leifum í kólesteról estera, sem byrja að mynda kjarnann og umbreytir nýmynduðu discoid HDL agnum í þroskaða kúlulaga HDL agnir. Þetta kólesterólríku lípóprótein er síðan skilað aftur í lifur, þar sem kólesteról er „losað“, sumt af þessu kólesteróli er breytt í gallsölt.
Mynd. 21-41. Viðbrögðin hvötuð af lesitín-kólesteróli-asýltransferasa (L CAT). Þetta ensím er til staðar á yfirborði HDL agna og er virkjað með apoA-1 (hluti af HDL brotinu). Kólesterólesterar safnast saman í nýstofnuðu HDL agnum og breyta þeim í þroskað HDL.

HDL er hægt að frásogast í lifur með viðtaka-miðluðum endocytosis, en að minnsta kosti er hluti HDL-kólesterólsins skilað í aðra vefi með öðrum aðferðum. HDL agnir geta bindst við SR - BI viðtakaprótein á plasmahimnu lifrarfrumna og í steraógeni eins og nýrnahettum. Þessir viðtakar miðla ekki endocytosis, heldur að hluta og sértækum flutningi kólesteróls og annarra lípíða HDL-hlutans inn í frumuna. Hið „tæma“ HDL brot fer síðan aftur í blóðrásina, þar sem það inniheldur nýja hluti af lípíðum úr chylomicrons og VLDL leifum. Sami HDL getur einnig fangað kólesteról sem er geymt í vefjum utan lifrar og flutt það í lifur með öfugt kólesterólflutning (Mynd 21-40). Í einu af öfugum flutningsafbrigðum hefst samspil HDL sem myndast við SR-BI viðtaka í kólesterólríkum frumum óvirkan dreifingu kólesteróls frá frumuyfirborðinu í HDL agnir sem flytja síðan kólesteról aftur í lifur. Í öðru afbrigði af öfugum flutningi í ríkri kólesterólfrumu, eftir klipping á HDL, hefur samskipti apoA-I við virka flutningsmanninn, ABC prótein. ApoA-I (og væntanlega HDL) frásogast með endocytosis, seytist síðan aftur, hlaðinn með kólesteróli, sem er fluttur í lifur.
Prótein ABC1 er hluti af stórri fjölskyldu flutningsaðila margra lyfja, þessir flutningsaðilar eru stundum kallaðir ABC flutningsaðilar, þar sem þeir allir innihalda ATP-bindandi snældur (ATP - bindandi snældur), þeir hafa einnig tvö himnulén með sex himnuflæði (sjá kafla. 11, v. 1). Þessi prótein flytja virkan mörg jón, amínósýrur, vítamín, sterahormón og gallsölt um himnur. Annar fulltrúi þessarar burðarfjölskyldu er CFTR prótein, sem með slímseigjusjúkdóm er skemmt (sjá viðbót 11-3, v. 1).
Kólesterólesterar fara inn í frumuna með viðtaka-miðluðum endocytosis
Hver LDL ögn í blóðrásinni inniheldur apoB-100, sem er þekkt af sérstökum yfirborðs viðtakapróteinum -LDL viðtaka á himnu frumna sem þurfa að ná kólesteróli. Binding LDL við LDL viðtakann setur af stað endocytosis, vegna þess sem LDL og viðtakinn hans fara inn í frumuna inni í endosominu (mynd 21-42). Endosominn smelst að lokum við lýsósómið, sem inniheldur ensím sem vatnsroða kólesterólester, sem losar kólesteról og fitusýrur í frumuefnið. ApoB-100 frá LDL brotnar einnig niður og myndar amínósýrur sem seytast út í cýtósólið, en LDL viðtakinn forðast niðurbrot og snýr aftur til frumuyfirborðsins til að taka aftur þátt í LDL upptöku. ApoB-100 er einnig til staðar í VLDL, en viðtaka-bindandi lén þess er ekki hægt að bindast við LDL viðtakann, umbreyting VLDLP í LDL gerir viðtakabindandi lénið í apoB-100 aðgengilegt. Michael Brown og Joseph Goldstein hafa rannsakað þessa kólesterólflutningaleið og viðtaka-miðluða endocytosis í markvef.
Michael Brown og Joseph Goldstein

Mynd. 21-42. Handtaka kólesteróls með viðtaka-miðluðum endocytosis.

Kólesteról, sem fer inn í frumurnar á þennan hátt, er hægt að fella í himnur eða estera það með ACAT (mynd 21-38) til geymslu í cýtósóli inni í lípíðdropum. Þegar nóg kólesteról er til staðar í LDL broti blóðs er komið í veg fyrir uppsöfnun umframmagns innanfrumukólesteróls með því að draga úr hraða myndunar þess.
LDL viðtakinn binst einnig við apoE og gegnir mikilvægu hlutverki í upptöku chylomicrons og VLDL leifa í lifur. Hins vegar, ef LDL viðtökur eru ekki fáanlegar (eins og til dæmis í músastofni með vantar LDL viðtaka gen), frásogast VLDL leifar og kýlómíkrónar enn í lifur, þó að LDL frásogist ekki. Þetta gefur til kynna tilvist hjálparforðakerfis fyrir viðtaka-miðluða endocytosis af VLDL og chylomicron leifum. Einn af viðtakaviðtakunum er LRP próteinið (lípóprótein viðtaka prótein), sem er tengt við lípóprótein viðtaka, sem binst apoE og fjölda annarra bindla.
Nokkur stig stjórnunar á nýmyndun kólesteróls
Kólesterólmyndun er flókið og ötullt dýrt ferli, svo það er greinilegt að líkaminn er hagstæður að hafa búnað til að stjórna nýmyndun kólesteróls, sem endurnýjar magn hans til viðbótar því sem fylgir matnum. Hjá spendýrum er kólesterólframleiðsla stjórnað af innanfrumuþéttni
kólesteról og hormón glúkagon og insúlín. Stig umbreytingar HMG - CoA í mevalonate (mynd 21-34) takmarkar hraðann í efnaskiptaferli kólesterólmyndunar (aðalatriðið í reglugerð). Þessi viðbrögð eru hvötuð af HMG - CoA redúktasa. Reglugerð til að bregðast við breytingum á kólesterólmagni er miðluð af glæsilegu eftirlitsstjórnunarkerfi fyrir gen sem kóðar HMG - CoA redúktasa. Þetta gen ásamt fleiri en 20 öðrum genum sem umrita ensím sem taka þátt í frásogi og nýmyndun kólesteróls og ómettaðra fitusýra er stjórnað af litlum fjölskyldu próteina sem kallast prótein sem hafa samskipti við sterólregluþáttinn í próteinmyndun (SREBP, bindandi prótein úr sterólreglum) . Eftir myndun eru þessi prótein kynnt í endoplasmic reticulum. Eina leysanlegi SREBP lénið með amínóstöðva virkar sem umritunarvirkjun með því að nota þá leið sem lýst er í kafla 10. 28 (v. 3). Hins vegar hefur þetta lén ekki aðgang að kjarnanum og getur ekki tekið þátt í virkjun gensins svo framarlega sem það er áfram í SREBP sameindinni. Til þess að virkja umritun HMG gensins - CoA redúktasa og önnur gen er aðskilnaðsvirka lénið aðskilið frá restinni af SREBP með prótýlýtískri klofnun. Þegar kólesteról er hátt eru SREBP prótein óvirk, fest á ER í fléttu með öðru próteini sem kallast SCAP (SREBP - virkjandi prótein fyrir klofun) (mynd 21-43). Það er SCAP sem bindur kólesteról og fjölda annarra steróla, sem starfa sem steról skynjari. Þegar magn steróls er hátt hefur SCAP - SREBP flókið líklega samskipti við annað prótein, sem heldur öllu fléttunni í ER. Þegar magn steróla í frumunni lækkar, leiðir sköpunarbreytingin á SCAP til þess að varðveislavirkni tapist og SCAP - SREBP fléttan flyst inn í blöðrurnar til Golgi fléttunnar. Í Golgi-fléttunni eru SREBP-prótein klofin tvisvar með tveimur mismunandi próteasum, önnur klofningin losar amínó-endalénið í cýtósólinu. Þetta lén færist til kjarnans og virkjar umritun markgena. SREBP prótein lénið með amínóstöðvum hefur stuttan helmingunartíma og brotnar hratt niður af próteasómum (sjá mynd 27-48, t. 3). Þegar sterólmagnið hækkar nægjanlega, er prótínsýting losunar á EB EBP prótein sviðum með amínóstöðvum aftur lokað og niðurbrot proteasome núverandi virkra léna leiðir til skjótrar lokunar markgenanna.
Mynd. 21-43. Virkjun SR EBP. SREB P prótein sem eru í samspili við sterólstýrt frumefni (grænn litur), strax eftir myndun, eru sett inn í ER og mynda flókið með S CAP (rauður litur). (N og C tákna amín og karboxýl enda próteina.) Í S-CAP bundnu ástandi eru SRE BP prótein óvirk. Þegar sterólmagnið lækkar, flyst SR EBP-S CAP flókið yfir í Golgi flókið, og SR EBP próteinin eru klofin í röð með tveimur mismunandi próteasum. Losaða amínósýra endanlegi SR EBP prótein lénið flytur yfir í kjarnann, þar sem það virkjar umritun stera-stýrðra gena.

Kólesterólmyndun er einnig stjórnað af nokkrum öðrum aðferðum (mynd 21-44). Hormónastjórnun er miðluð af samgildum breytingum á NM G-CoA redúktasa. Þetta ensím er til í fosfórýleruðum (óvirkum) og affosfórýleruðum (virkum) myndum. Glúkagon örvar fosfórýleringu (óvirkjun) ensímsins og insúlín stuðlar að fosfórýleringu, virkjar ensímið og styrkir nýmyndun kólesteróls. Hátt innanfrumuvökvaþéttni kólesteróls virkjar ASAT sem eykur estrunar kólesteróls til útfellingu. Að lokum hindrar mikið magn frumukólesteróls umritun gena sem umbreytir LDL viðtaka, dregur úr framleiðslu þessarar viðtaka og þess vegna upptöku kólesteróls úr blóði.
Mynd. 21-44. Reglugerð um kólesterólmagn veitir jafnvægi milli myndunar og frásogs kólesteróls úr mat. Glúkagon auðveldar fosfórýleringu (óvirkjun) af NM G-COA redúktasa, insúlín stuðlar að fosfórýleringu (virkjun). X - óþekkt umbrotsefni kólesteróls sem örva próteingreiningu á NM G -CoA redúktasa.

Óreglulegt kólesteról getur leitt til alvarlegra veikinda hjá mönnum. Þegar heildarmagn tilbúins kólesteróls og kólesteróls sem fæst úr fæðu fer yfir það magn sem þarf til himnusamsetningar, myndun gallsölt og sterum, getur sjúkleg uppsöfnun kólesteróls í æðum (æðakölkun) leitt til þess að þau eru stífluð (æðakölkun). Í iðnríkjum er það hjartabilun vegna hindrunar á kransæðum sem er aðalorsök dauðsfalla. Þróun æðakölkunar tengist miklu magni af kólesteróli í blóði og sérstaklega háu kólesteróli sem þolist af LDL brotinu, mikið gildi HDL í blóði, þvert á móti, hefur jákvæð áhrif á ástand æðanna.
Með arfgengri kólesterólhækkun (erfðagalla) er magn kólesteróls í blóði mjög hátt - alvarleg æðakölkun myndast hjá þessu fólki þegar á barnsaldri. Vegna gölluð LDL viðtaka kemur upp ófullnægjandi upptaka miðluð upptöku LDL kólesteróls. Fyrir vikið er kólesteról ekki fjarlægt úr blóðrásinni, það safnast upp og stuðlar að myndun æðakölkunarplata. Nýmyndun innræns kólesteróls heldur áfram, þrátt fyrir umfram kólesteról í blóði, þar sem utanfrumukólesteról kemst ekki inn í frumuna til að stjórna myndun innanfrumna (mynd 21-44).Til meðferðar á sjúklingum með arfgengan kólesterólhækkun og aðra sjúkdóma sem tengjast hækkuðu kólesteróli í sermi eru statín flokkar notaðir. Sum þeirra eru fengin úr náttúrulegum uppruna en önnur eru búin til af lyfjaiðnaðinum. Statín eru svipuð mevalonati (bæta við 21-3) og eru samkeppnishemlar NMS-CoA redúktasa.
Viðauki 21-3. LÆKNI. Lípíð tilgátan og stofnun statína
Kransæðahjartasjúkdómur (CHD) er aðalorsök dánartíðni í þróuðum löndum. Þrenging á kransæðum sem flytja blóð til hjarta á sér stað vegna myndunar fituflagna sem kallast æðakölkun (plaques); þessar veggskjöldur innihalda kólesteról, fibrillar prótein, kalsíum, blóðflögu blóðtappa og frumur brot. Á XX öld. Það var virk umræða um tengsl milli slagæðahindrunar (æðakölkun) og kólesteról í blóði. Þessar umræður og virkar rannsóknir í þessa átt hafa leitt til þess að árangursrík lyf hafa lækkað kólesteról.
Árið 1913 birti N.N. Anichkov, þekktur rússneskur vísindamaður og sérfræðingur á sviði tilrauna meinafræði, verk þar sem hann sýndi fram á að kanínur, sem eru fóðraðar með kólesterólríkan mat, mynda skemmdir á æðum sem líkjast gervigler í skipum aldraðra. Anichkov stundaði rannsóknir sínar í nokkra áratugi og birti niðurstöðurnar í þekktum vestrænum tímaritum. Því miður urðu gögn hans ekki grundvöllur fyrirmyndar fyrir þróun æðakölkun hjá mönnum þar sem á þeim tíma var tilgáta ríkjandi um að þessi sjúkdómur væri náttúrulegur afleiðing öldrunar og ekki sé hægt að koma í veg fyrir það. Hins vegar voru smám saman að safna gögnum um tengsl milli kólesteróls í sermi og þróunar æðakölkun (fitu tilgáta) og á sjöunda áratugnum. sumir vísindamenn hafa skýrt sagt að hægt sé að meðhöndla þennan sjúkdóm með lyfjum. Hins vegar var hið gagnstæða sjónarmið til þar til birt var árið 1984 niðurstöður breiðrar rannsóknar á hlutverki kólesteróls sem gerð var á vegum bandarísku heilbrigðisstofnunarinnar (Coronary Primary Prevention Trial). Sýnt var fram á tölfræðilega marktæka lækkun á tíðni hjartadreps og högg með lækkun kólesteróls í blóði. Í þessari rannsókn var kólesteról, anjónaskiptar plastefni sem bindur gallsýrur, notað til að lækka kólesteról. Niðurstöðurnar hafa örvað leitina að nýjum, öflugri meðferðarlyfjum. Ég verð að segja að í vísindaheiminum hvarf efasemdir um réttmæti fitu tilgátunnar alveg með tilkomu statína seint á níunda áratugnum - snemma á tíunda áratugnum.
Fyrsta statínið var uppgötvað af Akira Endo í Sankyo í Tókýó. Endo gaf út verk sín árið 1976, þó að hann hafi fjallað um vandamál kólesterólumbrots í nokkur ár. Árið 1971 lagði hann til að kólesterólmyndunarhemlar gætu einnig verið í sveppaframleiðendum sýklalyfja sem rannsökuð voru á þeim tíma. Í nokkur ár af mikilli vinnu greindi hann meira en 6.000 menningarheima af ýmsum sveppum þar til hann komst að jákvæðri niðurstöðu. Efnasambandið sem myndaðist var kallað samningur. Þetta efni lækkaði kólesteról hjá hundum og öpum. Þessar rannsóknir vöktu athygli Michael Brown og Joseph Goldstein frá Southwestern Medical School í Texas. Brown og Goldstein ásamt Endo hófu sameiginlega rannsókn og staðfestu gögn hans. Helstu velgengni fyrstu klínísku rannsóknarinnar tóku lyfjafyrirtæki við þróun þessara nýju lyfja. Hjá Merck hóf teymi undir forystu Alfred Alberts og Roy Wagelos nýja skimun á svepparækt og uppgötvaði samtals 18 menningu annað virkt lyf. Nýja efnið er kallað lovastatin. Samtímis var það almennt talið að gjöf stórra skammta af kompaktíni til hunda leiði til þróunar krabbameins og að leit að nýjum statínum á níunda áratugnum. hefur verið lokað. En á þeim tíma var ávinningur þess að nota statín til að meðhöndla sjúklinga með fjölskyldumeðferð með kólesterólhækkun þegar í ljós. Eftir fjölmargt samráð við alþjóðlega sérfræðinga og Matvæla- og lyfjaeftirlitið (FDA, Bandaríkin), byrjaði Merck að þróa lovastatin. Viðamiklar rannsóknir næstu tvo áratugi hafa ekki leitt í ljós krabbameinsvaldandi áhrif lovastatíns og nýju kynslóðar lyfja sem birtust í kjölfar þess.
Mynd. 1. Statín eru hemlar á NM G-CoA redúktasa. Samanburður á uppbyggingu mevalonats og fjögurra lyfja (statína) sem hindra verkun NM G -CoA redúktasa.

Statín hindra verkun HMG - CoA - redúktasa, líkja eftir uppbyggingu mevalonats og hindra þannig myndun kólesteróls. Hjá sjúklingum með kólesterólhækkun sem stafar af göllum í einu eintaki af LDL viðtaka geninu, þegar þeir taka lovastatin, lækkar kólesterólmagnið um 30%. Lyfið er jafnvel virkara í samsettri meðferð með sérstökum kvoða sem bindur gallsýrur og kemur í veg fyrir frásog frá þeim í þörmum.
Eins og er eru statín oftast notuð til að lækka kólesteról í blóði. Þegar einhver lyf eru tekin vaknar spurningin um óæskilega aukaverkanir þeirra. Hins vegar, þegar um er að ræða statín, eru margar aukaverkanir, aftur á móti, jákvæðar. Þessi lyf geta örvað blóðflæði, lagað nú þegar gervigúða (svo að þau brjótast ekki frá veggjum æðar og trufla ekki blóðflæði), hindra samloðun blóðflagna og einnig veikja bólguferli í veggjum æðum. Hjá sjúklingum sem taka statín í fyrsta skipti birtast þessi áhrif jafnvel áður en kólesterólmagn byrjar að lækka og tengist hugsanlega hömlun á ísóprenóíðmyndun. Auðvitað eru ekki allar aukaverkanir statína gagnlegar. Hjá sumum sjúklingum (venjulega meðal þeirra sem taka statín ásamt öðrum lyfjum sem lækka kólesteról) geta komið fram vöðvaverkir og vöðvaslappleiki og stundum í nokkuð sterku formi. Aðrar nokkuð margar aukaverkanir statína eru einnig skráðar, sem sem betur fer koma sjaldan fram. Hjá langflestum sjúklingum getur tekið statín hindrað þróun hjarta- og æðasjúkdóma. Eins og við á um öll önnur lyf, á aðeins að nota statín eins og læknirinn mælir með.

Með arfgengri fjarveru HDL kólesteróls er kólesterólmagn mjög lágt, með Tangier-sjúkdómi er kólesteról nánast ekki ákvarðað. Báðir erfðasjúkdómar stafar af stökkbreytingum í ABC1 próteini. HDL-lausa kólesterólhlutinn getur ekki fangað kólesteról úr frumum með skort á ABC1 og frumur sem tæma kólesteról eru fljótt fjarlægðar úr blóðinu og eytt. Bæði arfgeng fjarvera HDL og Tangier sjúkdóms eru mjög sjaldgæf (færri en 100 fjölskyldur með Tangier sjúkdóm eru þekktar um allan heim), en þessir sjúkdómar sýna hlutverk ABC1 próteins við að stjórna HDL plasmaþéttni. Þar sem lágt HDL gildi í plasma er í samræmi við mikið hlutfall af skemmdum á kransæðum, getur ABC1 prótein verið gagnlegt markmið fyrir lyf sem eru hönnuð til að stjórna HDL stigum. ■
Sterahormón myndast með því að kljúfa hliðarkeðju kólesterólsins og oxun þess.
Maður fær öll sterahormónin sín úr kólesteróli (mynd 21-45). Tveir flokkar sterahormóna eru búnir til í nýrnahettubarkar: steinefni í barksterum,sem stjórna frásogi ólífrænna jóna (Na +, C1 - og HCO) 3 -) í nýrum, og sykurstera, sem hjálpa til við að stjórna glúkógenógenmyndun og draga úr bólgusvörun. Kynhormón eru framleidd í æxlunarfrumum karla og kvenna og í fylgjunni. Meðal þeirra prógesterón sem stjórnar æxlunarferli kvenna, andrógen (t.d. testósterón) og estrógen (estradiol), sem hafa áhrif á þróun auka kynferðislegra einkenna karla og kvenna. Sterahormón hafa áhrif við mjög lágan styrk og eru því búnir til í tiltölulega litlu magni. Í samanburði við gallsölt er tiltölulega lítið kólesteról neytt til framleiðslu á sterahormónum.

Mynd. 21-45. Sum sterahormón myndast úr kólesteróli. Uppbygging sumra þessara efnasambanda er sýnd á mynd. 10-19, v. 1.

Til að mynda sterahormóna þarf að fjarlægja nokkur eða öll kolefnisatóm í „hliðarkeðju“ C-17 D-hringsins í kólesteróli. Fjarlæging á hliðarkeðju á sér stað í hvatberum stera af völdum vefja. Fjarlægingarferlið samanstendur af hýdroxýleringu af tveimur aðliggjandi kolefnisatómum hliðarkeðjunnar (C-20 og C-22), síðan klofning tengisins á milli þeirra (mynd 21-46). Myndun ýmissa hormóna felur einnig í sér upptöku súrefnisatóma. Öll hýdroxýlering og oxunarviðbrögð við myndun stera eru hvött með oxíðasa með blönduðum virkni (bæta við 21-1) sem nota NА D PH, O 2 og hvatbera cýtókróm P-450.
Mynd. 21-46. Klofningur á hliðarkeðjunni við myndun sterahormóna. Í þessu oxíðasa kerfi með blönduð virkni sem oxar aðliggjandi kolefnisatóm, virkar cýtókróm P-450 sem rafeindabær. Einnig taka þátt í ferlinu rafeindaflutandi prótein, adrenodoxin og adrenodoxin reductase. Þetta kerfi til að kljúfa hliðarkeðju fannst í hvatberum í nýrnahettum, þar sem virk framleiðsla á sterum fer fram. Pregnenolone er undanfari allra stera hormóna (mynd 21-45).

Milliverkanir á lífmyndun kólesteróls taka þátt í mörgum öðrum efnaskiptaferlum.
Til viðbótar við hlutverk sitt sem milliefni kólesteróllífsmyndunar, þjónar ísópentenýl pýrofosfat sem virkur undanfari við myndun gríðarlegs fjölda lífefnasameinda sem gegna ýmsum líffræðilegum aðgerðum (mynd 21-47). Má þar nefna A-, E- og K-vítamín, plöntulitun eins og karótín og blaðgrænu plöntukeðjuna, náttúrulegt gúmmí, margar ilmkjarnaolíur (til dæmis ilmandi grunnur af sítrónuolíu, tröllatré, moskus), ungum skordýrahormóni sem stjórnar myndbreytingu, dolichols, sem þjóna sem lípíðleysanleg burðarefni við flókna nýmyndun fjölsykríða, ubikínóns og plastókínínón - rafeindabærna í hvatberum og klórplastum. Allar þessar sameindir eru ísóprenóíð í byggingu. Meira en 20.000 mismunandi ísóprenóar hafa fundist í náttúrunni og greint er frá hundruðum nýrra á hverju ári.
Mynd. 21-47. Heildarmyndin á lífmyndun ísóprenóíða. Skipulag flestra endafurða sem kynntar eru hér eru gefnar í kafla. 10 (v. 1).

Prenýlering (samgild tenging við ísóprenóíð, sjá mynd 27-35) er algengur búnaður sem prótein festast við innra yfirborð frumuhimna spendýra (sjá mynd 11-14). Í sumum próteinum er bundið lípíð táknað með 15 kolefnis farnesýl hópi, í öðrum er það 20 kolefnis geranyl geranyl hópur. Þessar tvær tegundir af lípíðum festa mismunandi ensím. Hugsanlegt er að prenýleringuviðbrögð beini próteinum í mismunandi himnur eftir því hvaða lípíð er fest. Útbreiðsla próteina er annað mikilvægt hlutverk fyrir ísóprenafleiður - þátttakendur í efnaskiptaferli kólesterólsins.
Samantekt á kafla 21.4 Biosynthesis of cholesterol, steroids and isoprenoids
■ Kólesteról myndast úr asetýl-CoA í flókinni viðbragðsröð gegnum milliefni eins og β-hýdroxý-ß-metýlglutaryl-CoA, mevalonat, tvö virkjuð ísópren dimetýlallyl pýrofosfat og ísó-pentýl pýrofosfat. Þétting ísópren eininga gefur ósíklísk skvalen sem hjólar til að mynda þéttan hringkerfi og stera hliðarkeðju.
■ Nýmyndun kólesteróls er undir hormónastjórnun og að auki er það hindrað með því að auka styrk innanfrumuvökva, sem á sér stað með samgildum breytingum og stjórnun uppskriftar.
■ Kólesteról og kólesterólesterar eru fluttir af blóði sem blóðfituprótein. VLDL-hlutinn flytur kólesteról, kólesterólesterar og tríasýlglýseról úr lifrinni í aðra vefi, þar sem þríasýlglýserólunum er klofið með lípóprótein lípasa og VLDL er breytt í LDL. LDL-hlutinn, sem auðgaður er með kólesteróli og kólesterólesterum, er óbeint tekinn af viðtaka með endocytosis, en B-100 apolipoprotein í LDL er viðurkennt af himnaviðtaka. HDL fjarlægir kólesteról úr blóði og flytur það í lifur. Næringarskilyrði eða erfðagallar í umbroti kólesteróls geta leitt til æðakölkun og hjartadrep.
■ Sterahormón (sykurstera, steinefni og kynhormón) myndast úr kólesteróli með því að breyta hliðarkeðjunni og setja súrefnisatóm í sterakerfi hringanna. Mörg önnur ísóprenóíð efnasambönd eru framleidd úr mevalonati með þéttingu isopentenýl pýrófosfats og dímetýlallyl pýrofosfats ásamt kólesteróli.
■ Forforritun tiltekinna próteina beinir þeim að bindistöðum með frumuhimnum og er mikilvæg fyrir líffræðilega virkni þeirra.
Spurning 48. Reglu umbrot há fitusýra (ß-oxun og lífmyndun). Tilmyndun malonyl CoA. Asetýl CoA karboxýlasa, stjórnun virkni þess. Flutningur acyl Co-a um innri himnu hvatbera.
Aðal
magn fenýlalaníns er neytt
á tvo vegu:
kveikir
í íkornum,
snýr
í týrósíni.
Beygja
fenýlalanín til týrósíns fyrst og fremst
nauðsynlegt til að fjarlægja umfram
fenýlalanín, þar sem mikill styrkur er
Það er eitrað fyrir frumur. Menntun
týrósín skiptir ekki öllu máli
þar sem skortur á þessari amínósýru
í frumum gerist nánast ekki.
Aðal
fenýlalanín umbrot hefst
með hýdroxýleringu þess (mynd 9-29), í
sem leiðir til týrósíns.
Þessi viðbrögð eru hvötuð af sérstökum
monooxy-nase - fenýlalanín hydra (zsilase,
sem þjónar sem meðframleiðandi
tetrahýdróbíópterín (N4BP).
Virkni ensíma fer einnig eftir
nærveru Fe2.
Í
lifrin er fyrst og fremst hraðari virkjun
glýkógen (sjá kafla 7). Hins vegar hlutabréf
glýkógeni í lifrinni er eytt
18-24 klukkustundir í föstu. Aðalheimild
glúkósa þar sem birgðir nánast
glýkógen verður glúkónógenes,
sem byrjar að flýta í gegn
Mynd.
11-29. Miklar umbrotabreytingar
orka þegar skipt er um gleypið
ríki eftir aðsog. CT
- ketónlíkaminn, FA - fitusýrur.
4-6 klst
eftir síðustu máltíð. Undirlag
glýseról er notað við nýmyndun glúkósa,
amínósýrur og laktat. Í hávegum
myndun hraða glúkagonstyrks
fitusýrur minnkaðar vegna
fosfórun og óvirkjun
asetýl CoA karboxýlasa og hlutfall
p-oxun eykst. Hins vegar
aukið fituframboð til lifrarinnar
sýrur sem eru fluttar
frá fitugeymslum. Asetýl-CoA myndað
við oxun fitusýra er það notað
í lifur til myndunar ketónlíkama.
Í
fituvef með auknum styrk
glúkagon minnkaði myndunartíðni
TAG og fitusog er örvað. Örvun
fitusundrun - afleiðing örvunar
hormónaviðkvæmur TAG lípasi
adipocytes undir áhrifum glúkagon.
Fitusýrur verða mikilvægar
orkugjafa í lifur, vöðvum og
fituvef.
Svo
þannig á eftir frásogstímabilinu
blóðsykursstyrk er viðhaldið
á stiginu 80-100 mg / dl og magn fitunnar
sýrur og ketónlíkaminn eykst.
Sykur
sykursýki er sjúkdómur sem kemur upp
vegna algerra eða ættingja
insúlínskortur.
A.
Helstu klínískar tegundir sykurs
sykursýki
Samkvæmt
Alþjóðastofnunin
sykursýki í heilsugæslu
flokkað eftir mismun
erfðaþættir og klínískir
tvö meginform: sykursýki
Tegund I - insúlínháð (IDDM) og sykursýki
Tegund II - óháð insúlíni (NIDDM).
Reglugerð
myndun zhk Regulatory ensím
myndun lcd - asetýl CoA karboxýlasa.
Þetta ensím er stjórnað af nokkrum
leiðir.
Virkjun / aðgreining
ensímundireiningarfléttur. Í
óvirkt form asetýl CoA karboxýlasa
táknar aðskildar fléttur,
hver samanstendur af 4 undireiningum.
Virkjandi ensímsins er sítrat. Það örvar
sambland af fléttum, fyrir vikið
þar sem ensímvirkni eykst
. Inhibitor-palmitoyl-CoA. Hann hringir
flókin aðgreining og lækkun
ensímvirkni.
Fosfórýlering / úrfosfórun
asetýl CoA karboxýlasa. Í
eftir frásog eða í
líkamlegt starf glúkagonað
adrenalín í gegnum adenýlat sýklasa
kerfið er virkjað með prokinasa A og
örva fosfórýleringu undir einingum
asetýl CoA karboxýlasa. Fosfórýlerað
ensímið er óvirkt og myndun fitu
sýrur stöðvast.
Gleypið
tímabil insúlíns virkjar fosfatasa,
og asetýl-CoA karboxýlasa fer í
defosfórýlerað ástand. Síðan
undir áhrifum sítrats á sér stað
fjölliðun mótfrumanna ensímsins, og
hann verður virkur. Auk örvunar
ensím, sítrat framkvæmir annað
virka í myndun LCD. Gleypið
tímabil í hvatberum lifrarfrumna
safnast sítrat, þar sem
asýlleifin er flutt til
cýtósól.
Reglugerð
ß-oxunarhraði.
Β oxunar-efnaskiptaferli,
fast tengd vinnu CPE og almenns
leiðir til niðurbrots. Þess vegna hraði þess
stjórnað af frumuþörf fyrir
orka þ.e.a.s. með hlutföllum ATP / ADP og NADH / NAD, svo og viðbragðshraða CPE og
algeng leið til niðurbrots. Hraði
P-oxun í vefjum fer eftir framboði
undirlag, þ.e.a.s.
á magn fitunnar
sýrur sem fara inn í hvatbera.
Ókeypis fitusýrustig
í blóðinu rís við örvun
fitusog í fituvef meðan á föstu stendur
undir áhrifum glúkagons og meðan á líkamsrækt stendur
vinna undir áhrifum adrenalíns. Í þessum
fitusýrur verða
ríkjandi orkugjafi
fyrir vöðva og lifur, vegna
ß-oxun myndast með NADH og asetýl-CoA hamlandi
pyruvat dehýdrógenasa flókið.
Umbreyting myndun pyruvatts
frá glúkósa til asetýl-CoA hægir á sér.
Milli umbrotsefni safnast upp
glýkólýsu og einkum glúkósa-6-fosfat.
Glúkósa-6-fosfat hindrar hexokinasa
og aftra því
notkun glúkósa í ferlinu
glýkólýsa. Þess vegna er ráðandi
notkun lcd sem aðalheimildar
orka í vöðvavef og lifur
sparar glúkósa fyrir taugavef og
rauð blóðkorn.
Β oxunarhraði einnig
fer eftir virkni ensíma
karnitín asýltransferasa I.
Í lifrinni er þetta ensím hindrað.
malonyl CoA, efni sem myndast
með myndun lcd. Í frásogstímabilinu
glýkólýsa er virkjuð í lifur og
myndun asetýl-CoA eykst
frá pyruvat. Fyrsta myndun viðbrögð
LCD umbreytingu asetýl-CoA í malonýl-CoA.
Malonyl-CoA hindrar ß-oxun lcd,
sem hægt er að nota við myndun
feitur.
Menntun
malonyl-CoA frá asetýl-CoA reglugerðum
viðbrögð í lífmyndun lcd. Fyrstu viðbrögð
nýmyndun lcd umbreytingu asetýl-CoA í
malonyl CoA. Katalytískt ensím
þessi viðbrögð (asetýl Coa karboxýlasa),
tilheyra flokki ligasa. Hann hefur að geyma
kovalent bundið biotin. Í því fyrsta
co2 samgilt viðbragðsstig
binst biotin vegna orku
ATP, í 2. stigi COO- fluttur
á asetýl-CoA til að mynda malonýl-CoA.
Virkni asetýl CoA karboxýlasa
ákvarðar hraða allra síðari
myndun viðbragða lc
sítrat virkjar ensím í cýtósóli
asetýl CoA karboxýlasa. Malonyl CoA í
aftur á móti hamlar flutning hærra
fitusýrur frá cýtósóli í fylki
hvatbera hindrar virkni
ytri asetýl CoA: karnitín asýltransferasi,
þannig að slökkva á oxun hærri
fitusýrur.
Asetýl-CoA oxaloacetat →
HS-CoA sítrat

HSCOA ATP sítrat → asetýl-CoA ADP Pi oxaloacetat
Asetýl-CoA
í umfryminu þjónar sem upphafs undirlag
fyrir myndun lcd, og oxaloacetate í
cýtósól umbreytist árið
niðurstaðan sem pyruvat myndast.
Lífsgerving kólesteróls
Lífsgerving kólesteróls á sér stað í endoplasmic reticulum. Uppruni allra kolefnisatóma í sameindinni er asetýl-SCoA, sem kemur hingað frá hvatberum í sítrati, rétt eins og við myndun fitusýra. Lífsgerving kólesterólsins eyðir 18 ATP sameindir og 13 NADPH sameindir.
Myndun kólesteróls á sér stað í meira en 30 viðbrögðum, sem hægt er að flokka í nokkur stig.
1. Samsetning mevalonsýru.
Fyrstu tvö myndunarviðbrögðin falla saman við ketogenesisviðbrögðin, en eftir myndun 3-hýdroxý-3-metýlglutaryl-ScoA fer ensímið inn hýdroxýmetýl-glútarýl-ScoA redúktasa (HMG-SCOA redúktasi), myndar mevalonsýru.
 |